Что такое центрифугирование? Определение и принцип метода. Физико-химические методы исследований в цитологии
Лекция № 3.
Количество часов: 2
МЕТОДЫ ИССЛЕДОВАНИЯ КЛЕТОК
1. Световая микроскопия
2. Электронная микроскопия, п реимущества и недостатки. Разновидности электронной микроскопии
Клетки очень малы по размерам и в тоже время сложно устроены. Поэтому для успешного изучения строения и функционирования клетки необходимо знать и владеть соответствующими экспериментальными методами.
На первом этапе развития цитологии единственным способом изучения клетки было световое микроскопирование .
Микроскоп – это прибор, позволяющий получить увеличенное изображение мелких объектов, не видимых невооруженным глазом. В микроскопии принято использовать следующие единицы длины:
микрометр (1 мкм – 10 -6 м );
нанометр (1 нм – 10 -9 м );
ангстрем (1Å – 10 -10 м ).
Существуют световое и электронное микроскопирование. В световом микроскопе для получения увеличенного изображения используется свет, в электронном - поток электронов. Качество изображения определяется разрешающей способностью микроскопа. Разрешающая способность – это наименьшее расстояние, на котором оптика микроскопа может различить раздельно две близко расположенные точки. Разрешающая способность человеческого глаза составляет около 100 мкм. Это означает, что невооруженным глазом с расстояния 25 см наблюдатель со средней остротой зрения может отличить одну точку от другой, если они отстоят друг от друга на расстоянии не менее 100 мкм. Если рассматриваемые точки находятся на расстоянии менее 100 мкм, то они кажутся одной расплывчатой точкой. Лучший современный световой микроскоп дает возможность рассмотреть структуры с расстоянием между элементами около 0,25 мкм, электронный микроскоп - порядка 1,5 А.
Световое микроскопирование - это совокупность методов наблюдения микрообъектов с помощью различных оптических микроскопов. Эти методы существенно зависят от типа объектива микроскопа, вспомогательных приспособлений к нему, вида микрообъекта и способа подготовки его для наблюдения, а также от характера его освещения при наблюдении. Разрешающая способность светового микроскопа ограничена размерами, сравнимыми с длиной световой волны (0,4–0,7 мкм для видимого света). Однако многие элементы клеточной структуры значительно меньше по размерам. Кроме того, при использовании обычного светового микроскопа большинство структур живой клетки являются оптически пустыми. Оптически пустыми называют структуры, которые прозрачны и почти не отличаются по показателю преломления от окружающей их среды. Для выявления таких структур были разработаны различные способы фиксации и окраски материала.
Фиксация – это обработка, которая быстро прерывает процессы жизнедеятельности клетки и по мере возможности сохраняет неизменными структуру клеток и тканей. После фиксации клетки становятся проницаемыми для красителей, фиксируется местоположение и стабилизируется структура макромолекул.
Окрашивание применяется для оптической дифференциации клеточных структур, а также в цитохимических исследованиях для выявления мест локализации химических соединений. Например, основные красители (гематоксилин) обладают сродством к содержимому ядра, а кислотные красители (эозин) окрашивают цитоплазму. Для изучения живых клеток используются витальные (прижизненные) красители . Витальные красители сравнительно легко проникают в живые клетки и окрашивают некоторые структуры, не повреждая их. Все же витальные красители не совсем безвредны для клетки, и после длительного воздействия приводят ее к гибели. К витальным красителям относятся нейтральный красный (для окрашивания цитоплазмы), метиленовый синий (окраска комплекса Гольджи) и др. При помощи витальных красителей удалось доказать существование некоторых органоидов клетки, которых раньше принимали за артефакты.
Артефакт – изменение, которое возникает в ходе приготовления препарата.
Перед проведением исследований клетки или кусочки ткани обычно заливают в расплавленный парафин или специальную смолу. Использованная для заливки среда охлаждается или полимеризуется. В результате этого образуется твердый блок, который режут на очень тонкие срезы с помощью микротома. Обычно толщина срезов для световой микроскопии составляет 1-10 мкм. Недостатком этого метода является повреждение ряда структур клетки. Поэтому применяют метод приготовления срезов с помощью быстрого замораживания. Замороженную ткань режут на специальном микротоме (криотоме), оборудованном холодной камерой (криостатом).
Помимо обычной световой микроскопии изучение клетки проводится с помощью темнопольной, фазово-контрастной, флюоресцентной и некоторых других видов световой микроскопии.
Темнопольное микроскопирование. Темнопольный микроскоп в отличие от обычного снабжен специальным конденсором. В конденсоре имеется темная диафрагма, не пропускающая свет в центр поля зрения, так что объект освещается косым пучком. При этом в объектив микроскопа попадают только лучи, отраженные и рассеянные от поверхности объекта, что повышает контрастность некоторых структур и делает их видимыми. Темнопольную микроскопию применяют для наблюдения ряда структур в живой клетке. В частности темнопольную микроскопию используют для определения частоты повреждений акросомы у спермиев сельскохозяйственных животных.
Фазовоконтрастное микроскопирование. Фазовоконтрастный микроскоп был сконструирован Фрицем Зернике в 1932 г. Фазовоконтрастная микроскопия является отличным методом прижизненного наблюдения за клетками. Ее используют для изучения многих органоидов клетки и хромосом во время деления. В конденсоре фазовоконтрастного микроскопа имеется кольцевая диафрагма, через которую свет проходит в виде полого конуса, а остальные лучи поглощаются. В объективе находится фазовая пластинка, представляющая собой прозрачный диск, имеющий выемку. Форма и размер выемки совпадают с прямым изображением кольцевой диафрагмы. При помещении объекта между конденсором и объективом в задней фокальной плоскости объектива, кроме прямого изображения, появляется несколько перекрывающих друг друга дифракционных изображений диафрагмы. Выемка фазовой пластинки рассчитывается так, чтобы оба пучка лучей, образующих прямое и дифракционное изображение, отличались по оптическому пути на четверть длины волны. Таким образом, фазовые различия, которые раньше не улавливались глазом, превращаются в различия интенсивности и становятся видимыми.
Флуоресцентная микроскопия является хорошим методом прижизненного наблюдения клеток. Флуоресцентный микроскоп позволяет наблюдать флуоресценцию (свечение) ряда веществ и структур клетки. Флуоресценция объекта возбуждается ультрафиолетовыми или сине-фиолетовыми лучами от специальных источников света. Излучение объекта всегда имеет большую длину волны, чем возбуждающий свет. Объект рассматривается в лучах его флуоресценции, которые отделяются от лучей возбуждающего света при помощи светофильтров. Ряд веществ (некоторые витамины, пигменты, липиды) обладают собственной (первичной) флуоресценцией. Вещества клетки, не обладающие этим свойством, предварительно окрашивают специальными красителями – флюорохромами , а затем наблюдают вторичную флуоресценцию.
Электронное микроскопирование. В электронном микроскопе для построения изображения вместо света используют поток электронов в вакууме. Фокусировка электронного пучка производится не линзами, как в световом микроскопе, а электромагнитными полями. Изображение наблюдают на флюоресцирующем экране и фотографируют. Объекты при электронной микроскопии находятся в глубоком вакууме, поэтому предварительно их подвергают фиксации и специальной обработке. По этой причине с помощью электронного микроскопа можно изучать только убитые клетки. Кроме того, они должны быть очень тонкими, так как поток электронов сильно поглощается объектом. В этой связи в качестве объектов используют ультратонкие срезы толщиной 20-50 нм, помещенные на тончайшие пленки. В трансмиссионном (просвечивающем) электронном микроскопе электроны проходят сквозь объект так, как в световом микроскопе через него проходит свет. Просвечивающая электронная микроскопия применяется для изучения ультратонких срезов микробов, тканей, а также строения мелких объектов (вирусов, жгутиков и др.). В сканирующем электронном микроскопе точно сфокусированный пучок электронов движется взад и вперед по поверхности образца. При этом отраженные от его поверхности электроны собираются и формируют изображение. Преимущество использования этой разновидности электронного микроскопа заключается в том, что создается трехмерное изображение. Поэтому сканирующая электронная микроскопия используется для изучения поверхности объектов. Электронный микроскоп имеет разрешающую способность около 1–2 нм. Этого достаточно для изучения макромолекул.
Авторадиография. Этот метод основан на применении меченными радиоактивными изотопами веществ. Если добавить в среду радиоактивный изотоп, поглощаемый клетками в процессе метаболизма, то его внутриклеточную локализацию можно впоследствии выявить с помощью авторадиографии. При использовании этого метода тонкие срезы клеток помещают на пленку. Пленка темнеет под теми местами, где находятся радиоактивные изотопы. В качестве изотопов используют фосфор (P 32 ), железо (Fe 59 ), серу (S 35 ), углерод (С 14), тритий (H 3 ) и др.
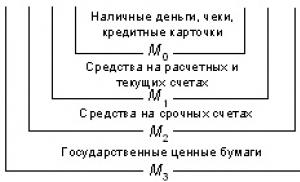
Центрифугирование. Начало методу было положено в 1926 г., когда Сведберг изобрел аналитическую центрифугу и использовал ее для определения молекулярной массы гемоглобина. Перед центрифугированием необходимо разрушить клеточную оболочку. Разрушение проводят, используя ультразвуковую вибрацию, осмотический шок, измельчение, продавливание через маленькое отверстие. При осторожном разрушении некоторые органоиды клетки сохраняются в интактном состоянии. Измельченные ткани с разрушенными клеточными оболочками помещают в пробирки и вращают в центрифуге с большой скоростью. Метод основан на том, что различные клеточные органоиды имеют разную массу и плотность. Более плотные органоиды осаждаются в пробирке при низких скоростях центрифугирования, менее плотные - при высоких. Эти слои изучают отдельно. Так ядра и неразрушенные клетки, быстро оседают при относительно низких скоростях и образуют осадок на дне центрифужной пробирки. При более высокой скорости выпадают в осадок митохондрии, а еще при более высоких скоростях и длительных периодах центрифугирования осаждаются рибосомы. Обычно такие очищенные компоненты сохраняют высокую биохимическую активность.
Метод культуры клеток и тканей состоит в том, что из одной или нескольких клеток на специальной питательной среде можно получить группу однотипных клеток. Этот метод имеет колоссальные перспективы не только для цитологии, но и для медицины, сельского хозяйства. Так клеточные культуры используют для выяснения закономерностей дифференцировки, взаимодействия клеток со средой, адаптации, старения, трансформации и др. В биотехнологии клеточные культуры применяют при производстве вакцин и биологически активных веществ. В фармакологии их используют в качестве тест-объектов при испытании новых лекарственных препаратов. Основоположником этого метода является американский зоолог и эмбриолог Р. Гаррисон (1879-1959), которому в 1907 г. удалось культивировать клетки саламандры в искусственной среде вне организма. Впоследствии многие типы растительных и животных клеток выращивались in vitro , и этот метод позволил сделать ряд важных открытий в области физиологии клеток. Выражение in vitro (по-латыни «в стекле) означает, что исследование проведено не на живом организме, а в стеклянном сосуде того или иного рода. В противоположность первому выражению in vivo указывает на эксперимент с целым, живым организмом. Культуры, приготовленные непосредственно из тканей организма, называют первичными культурами. В большинстве случаев клетки первичной культуры можно перенести из культуральной чашки и использовать для получения большого количества вторичных культур. Клеточные линии можно использовать для получения кло6нов, которые происходят из одиночной клетки-предшественника. Можно осуществить слияние клеток одного или разных видов. Чтобы добиться слияния, клетки подвергают воздействию вирусных ферментов или полиэтиленгликоля. Эти вещества повреждают плазматическую мембрану клеток, что приводит к образованию клетки с двумя отдельными ядрами. Спустя определенное время такая клетка делится путем митоза, образуя гибридную клетку. В гибридной клетке все хромосомы объединены в одно большое ядро. Такие гибридные клетки можно клонировать и получить гибридную клеточную линию. Используя этот метод, удалось получить гибридные клетки человека и мыши, человека и жабы. Поученные гибридные клетки нестабильны и после многочисленных клеточных делений теряют большинство хромосом либо одного, либо другого вида. Конечный продукт становится, например, по существу клеткой мыши, где человеческие гены отсутствуют или имеются лишь в незначительном количестве. Поэтому эту методику с успехом можно использовать для картирования генов в хромосомах человека.
Микрургия. Этот метод основан на использовании микроманипуляторов. Они представляют собой приборы, обеспечивающие точные движения микроинструментов в клетке. Микроинструменты обычно делают из стекла. Их форма определяется задачами микрургических операций. Они могут быть в виде игл, шприцев, пипеток, шпателей, скальпелей и т. д. С помощью микроманипуляторов над клетками можно производить разнообразные операции (инъекции в клетку веществ, извлечение и пересадка ядер, локальное повреждение клеточных структур и т.д.). Особенно хорошо микрургические операции удаются на крупных клетках (одноклеточные, яйцеклетки амфибий, клетки зародышей некоторых животных). Так клетку амебы удается разделить на три основных компонента – мембрану, цитоплазму и ядро. Затем эти компоненты можно вновь собрать и получить живую клетку. Таким путем могут быть получены искусственные клетки, состоящие из компонентов разных видов амеб. Микрургические операции производятся не только микроинструментами, но сфокусированным пучком ультрафиолетовых лучей (лучевой микроукол).
Кроме названных методов при изучении клетки используют хроматографию, электрофорез и некоторые другие. Новые методы позволили достичь огромных успехов в изучении клетки. Однако следует помнить, что классические методы цитологии, основанные на фиксации, окрашивании и изучении клеток под световым микроскопом, по-прежнему сохраняют практическое значение.
Лекция 1.
Структура растительной клетки
Световая микроскопия
Электронная микроскопия
Дифференциальное центрифугирование
Метод культуры клеток
Клетка является основной структурной и функциональной единицей живых организмов.
Клетки эмбриональных (неспециализированных) тканей животных и растений в общем плане строения очень сходны. Именно это обстоятельство в свое время явилось причиной для появления и развития клеточной теории. Морфологические различия проявляются уже в дифференцированных клетках специализированных тканей растений и животных. Особенности строения растительной клетки, как и растения в целом, связаны с образом жизни и способом питания. Большинство растений ведет относительно неподвижный (прикрепленный) образ жизни. Специфика питания растений состоит в том, что вода и питательные вещества: органические и неорганические, находятся вокруг в рассеянном виде и растению приходится их поглощать путем диффузии. Кроме того, зеленые растения на свету осуществляют автотрофный способ питания. Благодаря этому, эволюционно сложились некоторые специфические особенности строения и роста растительных клеток. К ним относятся:
| прочная полисахаридная клеточная стенка , окружающая клетку и составляющая жесткий каркас; |
|
| пластидная система , возникшая в связи с автотрофным типом питания; |
|
| вакуолярная система , которая в зрелых клетках обычно представлена крупной центральной вакуолью, занимающей до 95% объема клетки и играющей важную роль в поддержании тургорного давления ; |
|
| особый тип роста клеток путем растяжения (за счет увеличения объема вакуоли); |
|
| тотипотентность , то есть возможность регенерации полного растения из дифференцированной растительной клетки; |
|
| есть еще одна деталь, отличающая растительные клетки от клеток животных: у растений при делении клеток не выражены центриоли . |
Строение клетки в самом общем виде известно вам еще из курса общей биологии и при подготовке к вступительным экзаменам вы достаточно хорошо штудировали эту тему. Эта тема в разных аспектах рассматривается и в соответствующих университетских курсах (например, зоология беспозвоночных, низшие растения). Кроме того, более детальное знакомство с клеткой на высоком уровне предстоит в курсе "цитология". Нам же важно акцентировать внимание на специфических особенностях строения растительной клетки, причем преимущественно клетки высшего растения.
При самом поверхностном рассмотрении структуры типичной растительной клетки в ее составе обнаруживаются три основных компонента: (1) клеточная стенка , (2) вакуоль, занимающая в зрелых клетках центральное положение и заполняющая практически весь их объем и (3) протопласт , оттесняемый вакуолью к периферии в виде постенного слоя. Именно эти компоненты обнаруживаются на малом увеличении светового микроскопа. Причем клеточная оболочка и вакуоль являются продуктами жизнедеятельности протопласта.
Живое тело клетки? протопласт состоит из органоидов, погруженных в гиалоплазму . К организмам клетки относятся: ядро, пластиды, митохондрии, диктиосомы, эндоплазматический ретикулум, микротельца и др. Гиалоплазма с органеллами за вычетом ядра составляет цитоплазму клетки.
Для выражения размеров субклеточных структур используются определенные меры длины: микрометр и нанометр .
Микрометр в системе единиц измерения СИ величина, равная 10 -6 м . Говоря другими словами, микрометр (аббревиатура мкм) составляет 1/1000000 долю метра и 1/1000 долю миллиметра. 1 мкм = 10 -6 м . Старое название этой меры микрон .
Нанометр в той же системе представляет миллионную долю миллиметра 1 нм = 10 -9 м и тысячную долю микрометра.
Размеры и форма растительных клеток варьируются в широком диапазоне. В типичном случае размеры клеток высшего растения колеблются в пределах 10 - 300 мкм. Правда, встречаются клетки - гиганты, например, клетки сочной мякоти плодов цитрусовых составляют в поперечнике несколько миллиметров или чрезвычайно длинные лубяные волокна у крапивы достигают 80 мм длины при микроскопической толщине.
По форме различают изодиаметрические клетки, у которых линейные размеры во всех направлениях равны или отличаются незначительно (то есть длина, ширина и высота этих клеток сопоставимы). Такие клетки называют паренхимными (паренхима) .
Сильно вытянутые клетки, у которых длина во много раз (иногда в сотни и тысячи) превышает высоту и ширину, называют прозенхимными (прозенхима) .
Методы изучения растительной клетки
Для изучения клеток разработано и применяется множество методов, возможности которых определяют уровень наших знаний в этой области. Успехи в изучении биологии клетки, включая наиболее выдающиеся достижения последних лет, как правило, связаны с применением новых методов. Поэтому для более полного понимания клеточной биологии необходимо иметь хотя бы некоторое представление о соответствующих методах исследования клетки.
Световая микроскопия
Самым древним и, вместе с тем, наиболее распространенным методом изучения клетки является микроскопия. Можно сказать, что и начало изучения клетки было положено изобретением светового оптического микроскопа.
Невооруженный человеческий глаз имеет разрешающую способность около 1/10 мм. Это означает, что если вы смотрите на две линии, которые находятся друг от друга на расстоянии меньше 0,1 мм, они сливаются в одну. Чтобы различить структуры, расположенные более тесно, применяют оптические приборы, например, микроскоп.
Но возможности светового микроскопа не безграничны. Предел разрешения светового микроскопа задается длиной световой волны, то есть оптический микроскоп может быть использован только для изучения таких структур, минимальные размеры которых сопоставимы с длиной волны светового излучения. Лучший световой микроскоп имеет разрешающую способность около 0.2 мкм (или 200 нм), то есть примерно в 500 раз улучшает человеческий глаз. Теоретически построить световой микроскоп с большим разрешением невозможно.
Многие компоненты клетки близки по своей оптической плотности и без специальной обработки практически не видны в обычный световой микроскоп. Для того, чтобы сделать их видимыми, используют различные красители , обладающие определенной избирательностью.
В начале XIX в. Возникла потребность в красителях для окрашивания текстильных тканей, что в свою очередь вызвало ускоренное развитие органической химии. Оказалось, что некоторые из этих красителей окрашивают и биологические ткани и, что было уж совсем неожиданно, часто предпочтительно связываются с определенными компонентами клетки. Использование таких избирательных красителей дает возможность более тонко исследовать внутреннее строение клетки. Приведем лишь несколько примеров:
| краситель гематоксилин окрашивает некоторые компоненты ядра в синий или фиолетовый цвет; |
|
| после обработки последовательно флороглюцином и затем соляной кислотой одревесневшие оболочки клеток становятся вишнево - красными; |
|
| краситель судан III окращивает опробковевшие клеточные оболочки в розовый цвет; |
|
| слабый раствор йода в йодистом калии окрашивает крахмальные зерна в синий цвет. |
Для проведения микроскопических исследований большую часть тканей перед окраской фиксируют . После фиксации клетки становятся проницаемыми для красителей, а структура клетки стабилизируется. Одним из наиболее распространенных фиксаторов в ботанике является этиловый спирт.
Фиксация и окрашивание не единственные процедуры, используемые для приготовления препаратов. Толщина большинства тканей слишком велика, чтобы их сразу можно было наблюдать при высоком разрешении. Поэтому выполняют тонкие срезы на микротоме . В этом приборе использован принцип хлеборезки. Для растительных тканей изготавливают чуть более толстые срезы, чем для животных, поскольку клетки растений обычно крупнее. Толщина срезов растительных тканей для световой микроскопии около 10 мкм - 20 мкм. Некоторые ткани слишком мягкие, чтобы из них сразу же можно было получить срезы. Поэтому после фиксации их заливают в расплавленный парафин или специальную смолу, которые пропитывают всю ткань. После охлаждения образуется твердый блок, который затем режется на микротоме. Правда, для растительных тканей заливка применяется значительно реже, чем для животных. Это объясняется тем, что растительные клетки имеют прочные клеточные стенки, составляющие каркас ткани. Особенно прочны одревесневшие оболочки.
Однако заливка может нарушить структуру клетки, поэтому применяют еще и другой метод, где эта опасность уменьшена? быстрое замораживание. Здесь можно обойтись без фиксации и заливки. Замороженную ткань режут на специальном микротоме (криотоме).
Замороженные срезы, приготовленные таким способом, имеют явное преимущество, поскольку в них лучше сохраняются особенности естественной структуры. Однако их труднее готовить, а присутствие кристаллов льда все же нарушает некоторые детали.
Микроскопистов всегда беспокоила возможность потери и искажения некоторых компонентов клетки в процессе фиксации и окраски. Поэтому полученные результаты проверяют другими методами.
Весьма заманчивой представлялась возможность исследовать под микроскопом живые клетки, но так, чтобы более отчетливо проявились детали их строения. Такую возможность дают особые оптические системы: фазово-контрастный и интерференционный микроскопы. Хорошо известно, что световые волны, подобно волнам воды, могут интерферировать друг с другом, увеличивая или уменьшая амплитуду результирующих волн. В обычном микроскопе, проходя через отдельные компоненты клетки, световые волны меняют свою фазу, хотя человеческий глаз этих различий не улавливает. Но за счет интерференции можно преобразовать волны, и тогда разные компоненты клетки можно отличить друг от друга под микроскопом, не прибегая к окрашиванию. В этих микроскопах используют 2 пучка световых волн, которые взаимодействуют (налагаются) друг на друга, усиливая или уменьшая амплитуду волн, поступающих в глаз от разных компонентов клетки.
Электронная микроскопия
Возможности светового микроскопа, как уже было сказано, ограничиваются длиной волны видимого света. Его максимальная разрешающая способность составляет примерно 0.2 мкм.
Большой шаг вперед был сделан в микроскопии в 20-х годах нашего века, когда было обнаружено, что соответствующим образом подобранные электромагнитные поля можно использовать подобно линзам для фокусирования пучков электронов.
Длина волны электрона значительно меньше, чет длина волны видимого света, и если вместо света использовать электроны, то предел разрешения микроскопа может быть заметно снижен.
На основе всего этого был создан микроскоп, в котором вместо света используется пучок электронов. Первый электронный микроскоп сконструировали в 1931 г. Кнолл и Руска в Германии. Прошло, однако, много лет, прежде чем появилась возможность изучать при помощи этого микроскопа срезы тканей. Лишь в 50-е годы были разработаны методы изготовления срезов, обладающих необходимыми качествами. С этого времени началась новая эра микроскопии, и в науку буквально хлынул поток информации о тонком строении клеток (ультраструктуре клеток).
Сложности электронной микроскопии состоят в том, что для исследования биологических образцов необходима специальная обработка препаратов.
Первая трудность заключается в том, что электроны обладают очень ограниченной проникающей способностью, поэтому следует изготавливать ультратонкие срезы, толщиной 50 - 100 нм. Для того, чтобы получить столь тонкие срезы, ткани сперва пропитывают смолой: смола полимеризуется и формирует твердый пластмассовый блок. Затем с помощью острого стеклянного или алмазного ножа срезы нарезают на специальном микротоме.
Есть еще одна трудность: при прохождении через биологическую ткань электронов не получается контрастного изображения. Для того, чтобы получить контраст, тонкие срезы биологических образцов пропитывают солями тяжелых металлов.
Существует два основных типа электронных микроскопов. В трансмиссионном (просвечивающем) микроскопе пучок электронов, проходя сквозь специально подготовленный образец, оставляет его изображение на экране. Разрешающая способность современного трансмиссионного электронного микроскопа почти в 400 раз больше светового. Эти микроскопы имеют разрешающую способность около 0,5 нм (для сравнения: диаметр атома водорода около 0,1 нм).
Несмотря на столь высокое разрешение, просвечивающие электронные микроскопы имеют крупные недостатки:
Трехмерное (объемное) изображение получают с помощью сканирующего электронного микроскопа (ЭМ). Здесь луч не проходит через образец, а отражается от его поверхности.
Исследуемый образец фиксируют и высушивают, после чего покрывают тонким слоем металла? операция называется оттенением (образец оттеняют).
В сканирующем ЭМ сфокусированный электронный пучок направляется на образец (образец сканируют). В результате металлическая поверхность образца испускает вторичные электроны слабой энергии. Они регистрируются и преобразуются в изображение на телевизионном экране. Максимальное разрешение сканирующего микроскопа невелико, около 10 нм, но зато изображение получается объемным.
Метод замораживания-скалывания
Принципиально новые возможности электронной микроскопии открылись сравнительно недавно, после разработки метода "замораживания - скалывания". С помощью этого метода исследуются тончайшие детали строения клетки, при этом получается объемное изображение в трансмиссионном электронном микроскопе.
При обычном замораживании в клетках образуются кристаллики льда, которые заметно искажают их структуру. Во избежание этого клетки замораживают очень быстро при температуре жидкого азота (- 196 С). При таком мгновенном замораживании кристаллы льда не успевают образоваться, и клетка не испытывает деформаций.
Замороженный блок раскалывают лезвием ножа (отсюда и название метода). Затем, обычно в вакуумной камере, избыток льда удаляют возгонкой. Эта операция называется травлением. После травления более резко обозначается рельеф в плоскости скола. Полученный образец оттеняется, то есть на поверхность образца напыляется тонкий слой тяжелых металлов. Однако весь фокус состоит в том, что напыление производится под углом к поверхности образца. Это очень важный момент. Появляется эффект тени, изображение выглядит объемным.
В трансмиссионном микроскопе электронный луч способен проникнуть только через очень тонкие срезы. Обычная толщина оттененных образцов чрезмерно велика, поэтому органическую материю, подстилающую слой металла, необходимо растворить. В результате остается тонкая металлическая реплика (или отпечаток) с поверхности образца. Реплику и используют в трансмиссионном микроскопе.
Этот метод предоставил, например, уникальную возможность наблюдать внутреннее строение мембран клетки.
Дифференциальное центрифугирование
Помимо микроскопии, другим основным и широко распространенным методом изучения клеток является дифференциальное центрифугирование или фракционирование.
Принцип метода состоит в том, что при центрифугировании развивается центробежная сила, под воздействием которой взвешенные частицы оседают на дно центрифужной пробирки.
После того, как в начале 40-х годов начали использовать ультрацентрифугу, разделение клеточных компонентов стало вполне реальным.
Прежде, чем подвергнуть клетки центрифугированию, их необходимо разрушить - разрушить жесткий каркас клеточных оболочек. Для этого используют различные методы: ультразвуковую вибрацию, продавливание через маленькие отверстия или самое обычное измельчение растительных тканей пестиком в фарфоровой ступе. При осторожном применении методов разрушения можно сохранить некоторые органеллы целыми.
При высокоскоростном центрифугировании крупные компоненты клетки (например, ядра) быстро оседают (седиментируют), при относительно низких скоростях и образуют осадок на дне центрифужной пробирки. При более высоких скоростях в осадок выпадают более мелкие компоненты, такие как хлоропласты и митохондрии.
То есть при центрифугировании компоненты клетки распадаются на фракции: крупные и мелкие, поэтому второе название метода? фракционирование. При этом, чем выше скорость и длительность цетрифугирования, тем мельче полученная фракция.
Скорость седиментации (осаждения) компонентов выражается с помощью коэффициента седиментации, обозначаемого S.
Этапы дифференциального центрифугирования: низкая скорость (ядра, цитоскелет), средняя скорость (хлоропласты), высокая скорость (митохондрии, ризосомы, микротельца), очень высокая скорость (рибосомы).
Фракционированные клеточные экстракты, называемые также бесклеточными системами, широко используются для изучения внутриклеточных процессов. Только работая с бесклеточными экстрактами, можно установить детальный молекулярный механизм биологических процессов. Так, использование именно этого метода принесло триумфальный успех в изучении биосинтеза белка.
Ну и вообще, чистые фракции внутриклеточных структур можно подвергать любым видам анализа.
Метод культуры клеток
Клетки животных, выделенные в культуру (то есть помещенные на питательную среду), погибают после определенного числа делений, поэтому считаются трудным и неудобным объектом для культивирования. Другое дело клетки растений, способные делиться неограниченное число раз.
Метод культуры клеток облегчает изучение механизмов клеточной дифференциации у растений.
На питательной среде клетки растений образуют однородную недифференцированную клеточную массу? каллус. Каллус обрабатывают гормонами. Под влиянием гормонов клетки каллуса могут давать начало разным органам.
Строение и жизнедеятельность растительной клетки.
1. Строение растительной клетки: целлюлозная оболочка, плазматическая мембрана, цитоплазма с органоидами, ядро, вакуоли с клеточным соком. Наличие пластид - главная особенность растительной клетки.
2. Функции клеточной оболочки - придает клетке форму, защищает от факторов внешней среды.
3. Плазматическая мембрана - тонкая пленка, состоит из взаимодействующих молекул липидов и белков, отграничивает внутреннее содержимое от внешней среды, обеспечивает транспорт в клетку воды, минеральных и органических веществ путем осмоса и активного переноса, а также удаляет вредные продукты жизнедеятельности.
4. Цитоплазма - внутренняя полужидкая среда клетки, в которой расположено ядро и органоиды, обеспечивает связи между ними, участвует в основных процессах жизнедеятельности.
5. Эндоплазматическая сеть - сеть ветвящихся каналов в цитоплазме. Она участвует в синтезе белков, липидов и углеводов, в транспорте веществ. Рибосомы - тельца, расположенные на ЭПС или в цитоплазме, состоят из РНК и белка, участвуют в синтезе белка. ЭПС и рибосомы - единый аппарат синтеза и транспорта белков.
6. Митохондрии - органоиды, отграниченные от цитоплазмы двумя мембранами. В них с участием ферментов окисляются органические вещества и синтезируются молекулы АТФ. Увеличение поверхности внутренней мембраны, на которой расположены ферменты, за счет крист. АТФ - богатое энергией органическое вещество.
7. Пластиды (хлоропласты, лейкопласты, хромопласты), их содержание в клетке - главная особенность растительного организма. Хлоропласты - пластиды, содержащие зеленый пигмент хлорофилл, который поглощает энергию света и использует ее на синтез органических веществ из углекислого газа и воды. Отграничение хлоропластов от цитоплазмы двумя мембранами, многочисленные выросты - граны на внутренней мембране, в которых расположены молекулы хлорофилла и ферменты.
8. Комплекс Гольджи - система полостей, отграниченных от цитоплазмы мембраной. Накапливание в них белков, жиров и углеводов. Осуществление на мембранах синтеза жиров и углеводов.
9. Лизосомы - тельца, отграниченные от цитоплазмы одной мембраной. Содержащиеся в них ферменты ускоряют реакцию расщепления сложных молекул до простых: белков до аминокислот, сложных углеводов до простых, липидов до глицерина и жирных кислот, а также разрушают отмершие части клетки, целые клетки.
10. Вакуоли - полости в цитоплазме, заполненные клеточным соком, место накопления запасных питательных веществ, вредных веществ; они регулируют содержание воды в клетке.
11. Клеточные включения - капли и зерна запасных питательных веществ (белки, жиры и углеводы).
12. Ядро - главная часть клетки, покрытая снаружи двухмембранной, пронизанной порами ядерной оболочкой. Вещества поступают в ядро и удаляются из него через поры. Хромосомы - носители наследственной информации о признаках организма, основные структуры ядра, каждая из которых состоит из одной молекулы ДНК в соединении с белками. Ядро - место синтеза ДНК, иРНК, рРНК.
Дифференциальное центрифугирование
В случае дифференциального центрифугирования образцы центрифугируют определенное время при заданной скорости, после чего удаляют надосадочную жидкость. Этот метод полезен для разделения частиц, сильно различающихся по скорости седиментации. Например, центрифугирование в течение 5--10 мин при 3000-- 5000 д приводит к осаждению интактных бактериальных клеток, тогда как большинство клеточных фрагментов при этом остается в надосадочной жидкости. Фрагменты клеточной стенки и большие мембранные структуры можно осадить центрифугированием при 20 000--50 000 § в течение 20 мин, в то время как маленькие мембранные везикулы и рибосомы требуют для осаждения центрифугирования при 200 000 § в течение 1 ч.
Зональное центрифугирование
Зональное центрифугирование представляет собой эффективный способ разделения структур, имеющих сходную плавучую плотность, но различающихся по форме и массе частиц. В качестве примеров можно привести разделение субъединиц рибосом, различных классов полисом, а также молекул ДНК, имеющих различную форму. Центрифугирование осуществляют либо в бакет-роторах, либо в специально устроенных зональных роторах; для предотвращения конвекции при центрифугировании в стаканах бакет-ротора или в камере зонального ротора создают слабый градиент (обычно сахарозы). Пробу наносят в виде зоны или узкой полосы в самом верху градиентного столбика. Для субклеточных частиц обычно используется градиент сахарозы от 15 до 40% (вес/объем).
Этот метод основан на различиях в скоростях седиментации частиц, отличающихся друг от друга размерами и плотностью. Разделяемый материал, например гомогенат ткани, центрифугируют при ступенчатом увеличении центробежного ускорения, которое выбирается так, чтобы на каждом этапе на дно пробирки осаждалась определенная фракция. В конце каждой стадии осадок отделяют от надосадочной жидкости и несколько раз промывают, чтобы в конечном итоге получить чистую осадочную фракцию. К сожалению, получить абсолютно чистый осадок практически невозможно; чтобы понять, почему это происходит, обратимся к рассмотрению процесса, происходящего в центрифужной пробирке в начале каждой стадии центрифугирования.
Сначала все частицы гомогената распределены по объему центрифужной пробирки равномерно, поэтому получить чистые препараты осадков самых тяжелых частиц за один цикл центрифугирования невозможно: первый образовавшийся осадок содержит в основном самые тяжелые частицы, но, кроме этого, также некоторое количество всех исходных компонентов. Получить достаточно чистый препарат тяжелых частиц можно лишь при повторном суспендировании и центрифугировании исходного осадка. Дальнейшее центрифугирование супернатанта при последующем увеличении центробежного ускорения приводит к седиментации частиц средних размеров и плотности, а затем и к осаждению самых мелких частиц, имеющих наименьшую плотность. На рис. 2.3 изображена схема фракционирования гомогената печени крысы.
Дифференциальное центрифугирование является, по-видимому, самым распространенным методом выделения клеточных органелл из гомогенатов тканей. Наиболее успешно применяется этот метод для разделения таких клеточных органелл, которые значительно отличаются друг от друга по размерам и плотности. Но даже и в этом случае получаемые фракции никогда не бывают абсолютно гомогенными, и для их дальнейшего разделения применяют другие методы, описанные ниже. Эти методы, основанные на различиях в плотности органелл, обеспечивают более эффективное разделение благодаря тому, что центрифугирование осуществляют в растворах с непрерывным или ступенчатым градиентом плотности. Недостатком этих методов является то, что приходится тратить время на получение градиента плотности раствора.
Зонально-скоростное центрифугирование
Метод зонально-скоростного, или, как его еще называют, s-зонального центрифугирования, заключается в наслаивании исследуемого образца на поверхность раствора с непрерывным градиентом плотности. Затем образец центрифугируют до тех пор, пока частицы не распределятся вдоль градиента в виде дискретных зон или полос. Благодаря созданию градиента плотности удается избежать смешивания зон, возникающего в результате конвекции. Метод зонально-скоростного центрифугирования применяется для разделения гибридов РНК--ДНК, субъединиц рибосом и других клеточных компонентов.
Мы как раз начали применять еще новый в то время метод дифференциального центрифугирования. Он сводится к тому, что клетки разрушают в гомогенизаторе, а затем центрифугируют при последовательно возрастающих скоростях, после чего получают несколько фракций, состоящих из разных органелл (рис. 1.) Выделенные таким путем органеллы все еще сохраняют многие из своих функциональных свойств, которые можно затем изучить посредством биохимических методов. Наша задача состояла в том, чтобы установить местонахождение в этих фракциях некоторых ферментов, участвующих в обмене углеводов в печени крысы, и тем самым выяснить, с какими клеточными структурами связаны эти ферменты. Как правило, мы проверяли сначала гомогенат клеток на присутствие данного фермента, а затем искали его во фракциях. Среди прочих ферментов мы работали с так называемой кислой фосфатазой. Этот фермент, отщепляющий неорганический фосфат от ряда фосфорных эфиров, не связан непосредственно с углеводным обменом. В основном он служил нам в качестве контроля.
К нашему удивлению, активность кислой фосфатазы в гомогенате была в 10 раз меньше той величины, которую следовало ожидать на основании предыдущих анализов препаратов, подвергнутых более основательному разрушению в смесителе Уоринга. Суммарная активность всех фракций хотя и превышала вдвое активность гомогената, но все-таки была в 5 раз меньше ожидаемой величины. Когда через пять дней мы повторили определения на тех же фракциях (которые хранили все время в холодильнике), то оказалось, что активность фермента значительно возросла во всех отдельных фракциях, а особенно во фракции, содержащей митохондрии. Теперь суммарная активность уже достигла ожидаемой величины.
К счастью, мы устояли перед искушением отбросить первую серию данных как результат технической ошибки. Мы провели несколько дополнительных опытов и быстро получили ключ к разгадке тайны. В живых клетках фермент в основном (или даже полностью) заключен внутри небольших мешочкоподобных частиц. Поверхностная мембрана этихчастиц не только способна удерживать фермент внутри частицы, но и препятствует проникновению извне тех мелких молекул фосфорных эфиров, с которыми мы работали. Активность, измеренная в наших опытах, характеризовала лишь ту небольшую часть фермента, которая либо находилась в клетке в свободном состоянии, либо освободилась из частиц, поврежденных в ходе опыта. Смеситель Уоринга практически разрушает все частицы; при более мягкой обработке в гомогенизаторе, которую мы применили в наших исследованиях, разрушается лишь около 10% частиц. Этим и объясняется низкая первоначальная активность фермента в гомогенате. Дальнейшее фракционирование приводит к возрастанию суммарной активности во фракциях еще на 10%. Остальная часть фермента освобождается в результате старения частиц при хранении их в течение пяти дней в холодильнике.
TBegin--> TEnd-->
TEnd-->
Рис. 1. Дифференциальное центрифугирование позволяет разделить клетки на фракции, состоящие из различных клеточных компонентов. Быстрое механическое вращение пестика разрушает клетки, вызывая освобождение их содержимого в окружающую среду. Гомогенат подвергается последовательному центрифугированию при разных скоростях. Метод, разработанный В. Шнайдером, включает этапы 1—8. Автор и его сотрудники ввели этапы 9 и 10. Фракция митохондрий (6 этап) осаждается после центрифугирования при 25 000 g в течение 10 мин. Числа, характеризующие градиент плотности сахарозы, показывают удельный вес (г/см3).
Пояснение.
Гипотеза, предполагающая, что сходство некоторых мух с пчёлами защищает их от врагов, проверяется экспериментально
Ответ: 2
Можно ли считать простым совпадением то, что эти мухи отличаются от своих близких родичей-комнатных мух-и похожи на неродственных им пчел? Современные биологи усомнятся в этом и выдвинут гипотезу, что окраска этих мух возникла в процессе эволюции, поскольку сходство с пчелами давало им какое-то преимущество (изучением эволюции мы займемся в гл. 2). В чем может заключаться это преимущество?
Для того чтобы ответить на такой вопрос, нужны какие-то идеи или гипотезы, которые позволят объяснить сделанные наблюдения. Так, можно предположить, что сходство с пчелами защищает мух от поедания хищниками или же что оно позволяет мухам, обманув пчел, проникнуть в улей и полакомиться медом.
Следующий шаг состоит в том, чтобы разработать и провести эксперименты, которые позволят проверить выдвинутые гипотезы. Некоторые гипотезы для науки бесполезны, потому что их нельзя проверить. Например, гипотеза о том, что «хищники принимают безобидных мух за опасных пчел», не поддается проверке, так как вы не можете узнать, о чем думает то или иное животное. Но даже если гипотезу можно проверить, это обычно не удается сделать непосредственно; надо прежде придумать какое-нибудь вытекающее из нее и поддающееся проверке предсказание.
Допустим, что вы на основе своей первой гипотезы (о том, что окраска мухи защищает ее от хищников) делаете следующее предсказание: если данный хищник был ужален пчелами и научился не трогать их, то он не станет нападать на муху, которая выглядит как пчела. Это предсказание можно проверить.
Для проведения соответствующего эксперимента нужен хищник, питающийся насекомыми. Таким хищником может быть, например, жаба, которая ловит насекомых, летающих или ползающих поблизости от нее. Если в садок с «неопытной» жабой, ранее не встречавшейся с пчелами, выпустить этих насекомых, то она поймает несколько штук, поймет, что они жалят, и откажется от дальнейшей охоты. Если затем, поместив в садок муху, имеющую полосатую, черную с желтым, окраску, мы убедимся, что жаба откажется от этой мухи, то, следовательно, наша гипотеза, согласно которой сходство мухи с пчелой спасает ее от хищников, подтверждается.
Но, быть может, пчелы здесь совсем ни при чем. Быть может, жабы просто не едят полосатых мух. Чтобы проверить это, придется использовать еще одну «неопытную» жабу. Если окажется, что жаба охотно пожирает мух в черную и желтую полоску, то, значит, вы получили дополнительное подтверждение гипотезы о том, что преимущество полосатого наряда мухи объясняется его сходством с окраской пчелы.
Настоящий научный эксперимент должен непременно сопровождаться контрольным экспериментом, который отличался бы от основного эксперимента одним (и только одним) фактором. В рассматриваемом здесь случае необходимо, чтобы обе используемые жабы практически не отличались друг от друга, т.е. принадлежали к одному виду, были одного пола, одного возраста и имели одинаковые размеры. Их следует содержать в одинаковых садках, при одинаковых условиях освещения, температуры, влажности и т.п. Единственное различие между ними должно состоять в том, что одна жаба до того, как ей предложат муху, уже имела возможность столкнуться с пчелой, а другой жабе такой случай не представился. Без контрольного эксперимента может возникнуть мысль, что жаба отказывается от полосатых мух по какой- то иной, не учтенной нами причине, например потому, что она не голодна, вы же делаете ошибочное заключение, что дело здесь в сходстве с пчелами. (Фактически вы можете провести еще один контрольный эксперимент, предложив первой жабе безобидную комнатную муху после того, как она откажется от полосатой мухи; это позволит проверить, действительно ли жаба сыта.)
Итак, вы провели хорошо поставленный научный эксперимент. Какие выводы вы можете сделать? Вернемся снова к гипотезе: «сходство мухи с пчелами защищает ее от хищников». Удалось ли вам доказать это? Нет; вы всего лишь показали, что одна жаба отказалась от полосатой мухи после того, как она научилась отказываться от пчел, тогда как другая жаба, никогда не имевшая дела с пчелами, поедала полосатых мух.
Исследователи не принимают результаты эксперимента до тех пор, пока не убедятся в их воспроизводимости. Возможность многократного воспроизведения результатов позволяет избежать ошибок. Достоверность результатов можно повысить, если многократно повторить эксперимент, используя большое число жаб и в точности воспроизводя те же условия. Сколько следует использовать жаб? Чем больше, тем лучше, но было бы непрактично (и утомительно) проводить эксперимент на всех жабах земного шара. На самом деле существуют статистические методы, позволяющие определить, насколько «надежны» результаты при данной величине выборки.
Вы можете также подвергнуть проверке предсказание о том, что окраска мух отпугивает «знакомых с пчелами» хищников; для этого следует проделать дополнительные эксперименты, используя вместо жаб лягушек, птиц, ящериц и других хищников, питающихся насекомыми. Чем больше альтернативных гипотез вам удастся опровергнуть и чем большее число разных хищников откажется от полосатых мух после близкого знакомства с пчелами, тем убедительнее вы докажете правильность своей гипотезы.
Гипотеза, подтвержденная многочисленными и разнообразными данными, полученными в результате воспроизводимых экспериментов, обычно считается теорией и в конце концов рассматривается как научно установленный «факт».